The Triassic
Biostratigraphy and phylogenetic evolution
The Permian-Triassic transition represented a critical period in foraminiferal evolutionary history, resulting in the greatest reduction of biodiveristy in Earth 's history and accounting for the extinction of approximately 85% of all living genera (Erwin et al., 2002; BouDagher-Fadel, 2008). Fusulininina, which was the dominant suborder throughout the Paleozoic, was nearly wiped out completely. The suborder Lagenina was reduced by 30%. The suborder Miliolina was reduced by 50% and the suborder Textulariina was reduced by 80%.
The mass extinction perpetuated a distinct change in the taxonomic composition of the calcareous benthic assemblages (Groves and Altiner, 2004). During the Early Triassic, shallow marine foraminifera were simple and rare, with the first recorded forms appearing in the eastern
paleo-Tethyan realm (Sweet et al., 1992; Ma´rquez, 2005), resulting elsewhere in a taphonomically induced gap in the understanding of the transition of the Permian survivors (Groves, 2000). These "disaster forms" were initially dominated by small arenaceous forms (Tong and Shi, 2000). The larger foraminifera in the Early and Middle Triassic evolved slowly, and it was not till the Ladinian stage that highly diversified communities, such as the Ammodiscoidea and the Endothyroidea thrived. Towards the end of the Triassic a gradual decline of the larger foraminifera groups occurred, which climaxed with an end Triassic mass extinction event (Tanner et al., 2004), where most remaining larger foraminifera disappeared. The biostratigraphic range of the main Triassic forms are summarized in the figure below:
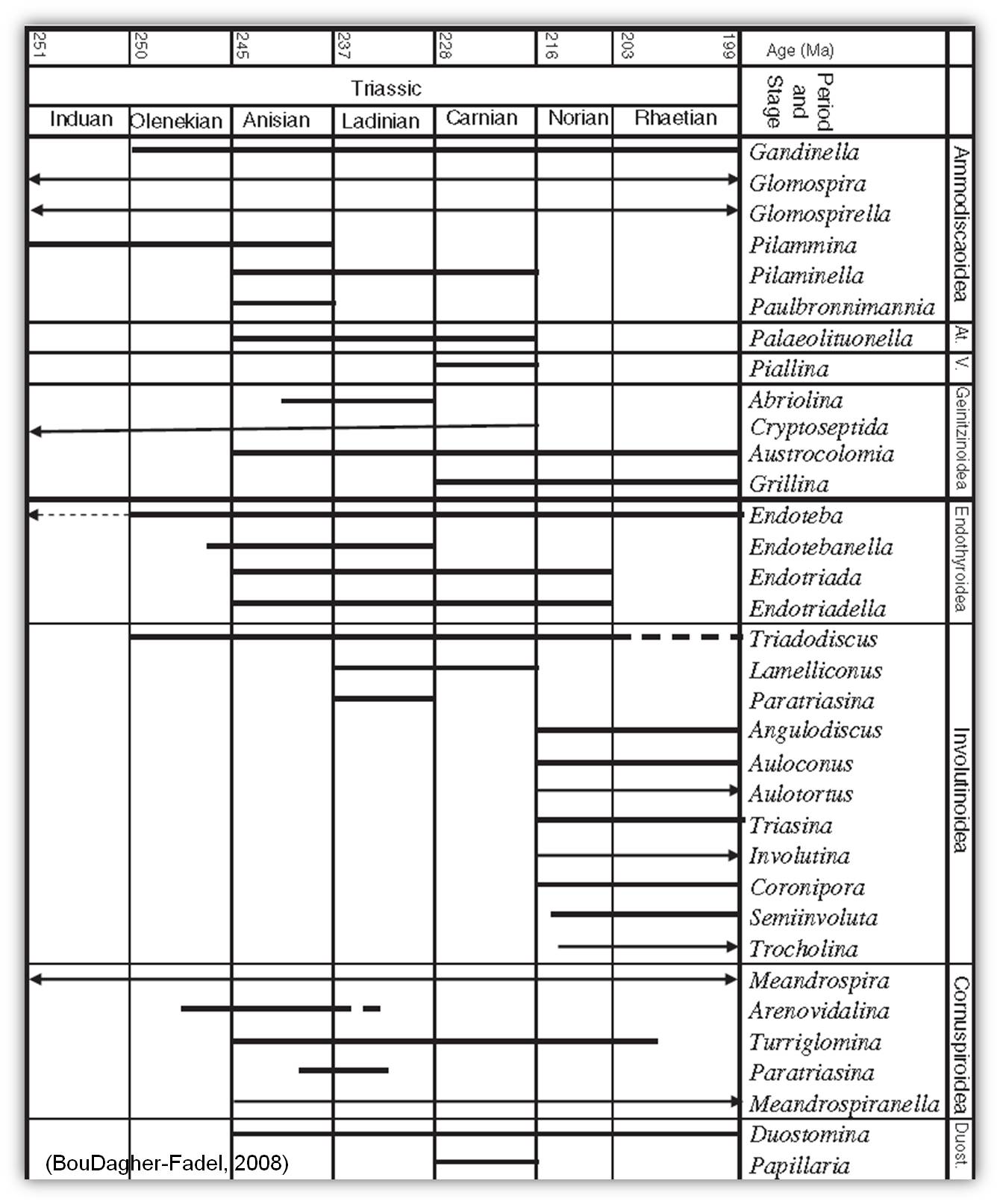
Biostratigraphic ranges of the main Triassic genera (At. = Ataxophragmiodea; V. = Verneuilinoidea)
Paleoecology of the Triassic large benthic foraminifera
During the Triassic, Pangea straddled the equator and the Tethys sea intruded Pangea from the east, resulting in a division between Laurasia in the north and Gondwana in the south. Sea levels were low with little or no dissolved oxygen and were possibly alkaline (Woods, 2005). Land animals migrated between the South and North Poles and the land was mostly dry and arid. This warm "super-hothouse" environment began during the Permian and may have contributed to the Permian-Triassic extinction due to high levels of carbon dioxide and low levels of oxygen (Berner et al., 2003; BouDagher-Fadel, 2008) shown on the figure below. The End Permian extinction led to a profound decline in reef production and caused the near-total elimination of metazoan reef systems (Hallam and Wignall, 1997), giving rise to the so-called reef gap (Fagerstrom, 1987; Flu¨ gel, 1994) during the Induan and Early Olenekian.
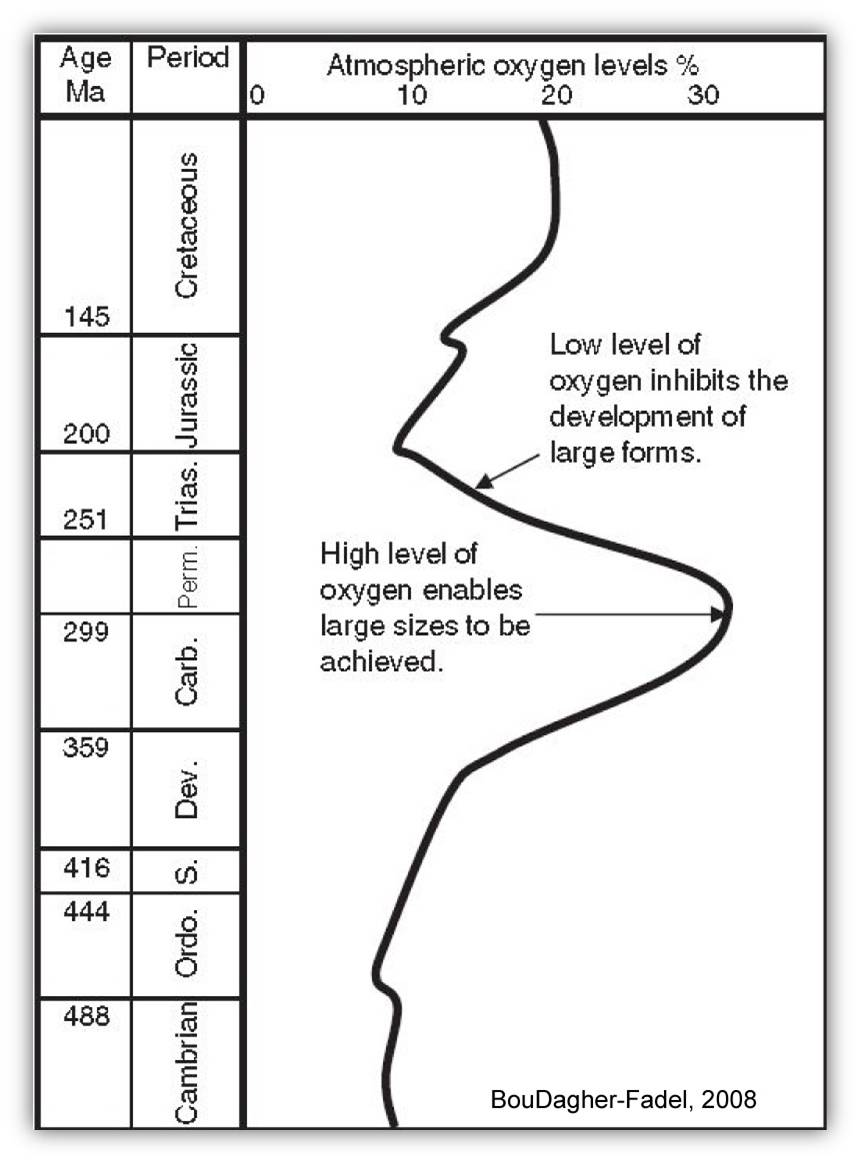
Oxygen variation over time with low levels noted during the Triassic (modified after Berner et al., 2003)
Recovery was very slow for foraminifera, and
ecological recovery was delayed until the Middle Olenekian. Many genera which seemed to have
disappeared at the Paleozoic–Mesozoic boundary remained hidden, possibly in
geographically restricted environments, before returning as ‘‘Lazarus’’ taxa in the
Olenekian or Middle to Late Triassic. It was not before the Anisian that larger benthic foraminifera became more widespread and cosmopolitan. During this time, a tropical sea formed between the
supercontinents of Gondwana (in the south) and Laurasia (in the north). This sea, called the Tethys, became home to large diverse assemblages of foraminifera. However, the larger Triassic benthic foraminifera never recovered the relatively gigantic sizes seen in the Permian, but rather remained small and nondistinct.
Anoxia has been regularly correlated with the dramatic Permian-Triassic
extinction events in the ocean
and the relatively small size of foraminifera during the Triassic (Erwin, 1993; Knoll
et al., 1996; Wignall and Twitchett, 1996). In the oceans today, many benthic
invertebrates compensate for hypoxic conditions via a broad spectrum of behavioral and morphological adaptations (Rogers, 2000).
During the Anisian,
carbonate platform development reoccurred with the important contribution of reefal debris, which was almost totally absent in Early Triassic
carbonate platforms. This created suitable habitats for the newly developing larger benthic foraminifera. New genera of arenaceous, fusulinine and milioline foraminifera colonized Tethys for the first time since the Permian-Triassic extinction (BouDagher-Fadel, 2008). During the Ladinian there was a clear stabilization of marine ecosystems, accompanied by the development of wide
carbonate platforms bearing highly diversified communities. This period saw the appearance of the first coral reefs formed by Scleractinia corals, calcareous algae and sponges. The end result is the appearance of much more stable and diverse living communities comprised of a large number of taxa (Ma´ rquez and Trifonova, 2000; Ma´ rquez, 2005).
The climate in the Carnian was unusually arid and hot and the seaway itself appears to have experienced a ‘‘salinity crisis’’ (general evaporation) and a breakdown of the reef system. In the Carnian, foraminiferal assemblages were dominated by fusulinines and miliolines. The Late Carnian–Norian interval saw major reef extinctions in the Tethys province, with the loss of older coral species, but diversity was maintained with reciprocal replacement by new taxa (Stanley, 2001). At this time the new foraminiferal taxa of
Involutinoidea became dominant and corals diversifed, constituting a new reef-building consortium. There was a period of worldwide expansion of
carbonate platforms and maximum reef diversity (Stanley, 2003). Associations of Late Triassic benthic foraminifera are generally typical of low energy, bay or lagoon-type, protected settings of salinity sometimes higher than normal, on shallow
carbonate ramps. A paleogeographic reconstruction by R. Blakey ( Ron Blakey and Colorado Plateau Geosystems Inc ™) of the earth during the Late Triassic is shown below.
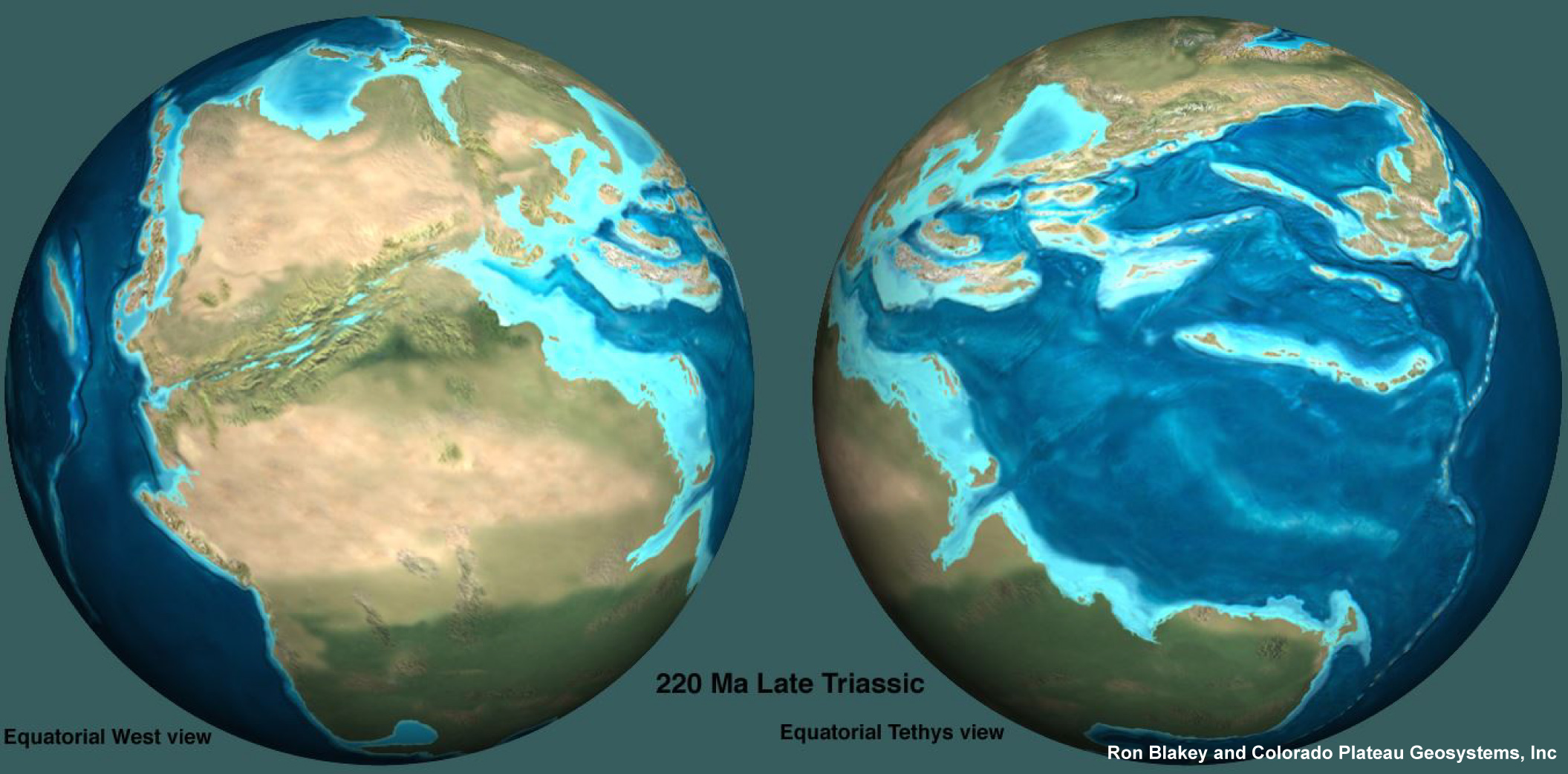
A paleogeographic reconstruction of the Late Triassic
(R. Blakey http://cpgeosystems.com/paleomaps.html)
Paleogeographic distribution of the Triassic large benthic foraminifera
The distribution of the Triassic genera are divided into three provinces: Europe and Russia, China and East Asia, and North America, and this distribution is shown in the figure left. There is a general and progressive increase in diversity from the Induan to the Anisian. The end of the Olenekian is marked by the increase in the number of suborders that exhibit larger morphologies. The ancestor of the Involutinoidea appeared in all the three provinces, and the group increased in diversity in China, East Asia and Europe during the Ladinian, but it was not before the Norian that they became widespread in Europe. The Fusulinoidea were common during the Anisian and Ladinian in China, East Asia and Europe, but they decreased gradually toward the end of the Norian. They did not disappear completely from these provinces till the end of the Rhaetian.
Throughout the Triassic, the diversity in North America was low, but there was a distinct increase in the number of genera throughout Europe, and to a lesser extent in China and East Asia, in the Anisian and Norian. There was a sharp decline in speciation during the Ladinian, and no new forms appeared at all in the Rhaetian. During the Anisian and the Norian, however, the larger benthic foraminifera speciation was at its most productive. North American foraminifera speciation mirrored generic diversity but remained poor throughout the Triassic.
There was a gradual increase in the number of extinctions from the Ladinian to the Carnian in all three provinces, to be followed by the largest extinction event at the end of the Rhaetian.
BouDagher-Fadel (2008) includes a brief but thorough analysis of the palaeogeography of the Triassic to explain the distributional trends seen during this time period and readers are referred to this work for more detail. After the great extinction at the Permian-Triassic boundary, foraminifera struggled to survive during the Induan interval. The recovery process was very slow in the Induan, but new forms were appearing, and, in fact, 80% of the Induan foraminifera were new Triassic forms. During the Early Triassic, the diversity of the foraminifera increased gradually in all three provinces, North America having the least diversity. This is probably due to its relative isolation during the Early Triassic (BouDagher-Fadel, 208). In the eastern Tethys area, the recovery process seems to be dominated by forms of the suborder Miliolina (Ma´ rquez, 2005).
During the Olenekian, Triassic foraminifera started quickly to diversify, leading to the highest number of new genera in the Anisian. At the end of the Anisian a small extinction occurred, mostly in the European Realm among the Ammodiscoidea (affecting 35% of the genera), and within the Ladinian, 60% of the Cornuspiroidea from all three provinces disappeared. This may have been associated with environmental stress caused by the large volcanic event (ca. 235 Ma) along the northwest margin of North America that formed the Wrangellia Terrane. After this event, the foraminifera in North America and Europe did not recover completely until the end of the Carnian. The Far East seems not to have been affected by this event.
Early Carnian Triassic foraminifera were cosmopolitan, as a small seaway connected the Tethys Ocean with North America (see Fig. 3.13). Foraminifera thrived until the end of the Carnian. The high number of extinctions at the end of the Carnian coincide with the occurrence of set of multiple impacts (Spray et al., 1998). Five terrestrial impact structures have been found to possess comparable ages
(214þ/�2 Ma), coincident with the Carnian–Norian boundary. These craters are Rochechouart (France), Manicouagan and Saint Martin (Canada), Obolon (Ukraine) and Red Wing (USA).
A map showing the position of the major, Late Carnian impact craters.
Soon though, foraminifera recovered and new forms filled the empty niches.The Norian saw the expansion of the
Involutinoidea in European Tethys. The Late Triassic also witnessed the appearance in the West Pangean reefs of reportedly Permian Lazarus taxa (Stanley, 2001). At the end of the Triassic, an abrupt extinction affected all groups in the marine realm and this extinction is one of the five largest
extinction events that took place during the Phanerozoic (Hallam, 1990). Only 16% of the Triassic foraminifera survived the End Triassic extinction. There has been much speculation
as to the cause of this
extinction event. Tanner et al. (2004) notes that the Late Triassic extinction appears protracted rather than catastrophic. Marine
regression reduced the available shallow marine
habitats for the Triassic foraminifera, and consequent competition may have been the forcing mechanism for extinction (Newell, 1967). Tucker and Benton (1982) specifically cited climate-induced changes occurring during the Late Triassic as a possible cause for mass extinction. Evidence of anoxia has also been documented in Triassic–Jurassic sections and has been suggested to be a significant factor in this event (Hallam and Wignall, 1997, 2000). However, the identification of clear evidence of iridium anomalies at the Triassic–Jurassic boundary (Tanner and Kyte, 2005) again raises the possibility that the mass extinction resulted from an extraterrestrial impact, or was triggered by flood
The synchronicity between Central Atlantic Magmatic Province (CAMP) volcanism and of marine faunal extinction at the Triassic–Jurassic boundary was recently confirmed by Schaltegger et al. (2007), who carried out biostratigraphic correlation between zonal ammonites and zircon ages. CAMP volcanism would have created anoxic and dysoxic environments caused by rising CO2 levels and related greenhouse effects during the rapid eruption of the flood
basalt.